Biomembran
| Forelder |
| celle |
| Underordnet |
|
Cellemembran Endomembransystem Ytre membran Dobbel membran |
| Genontologi |
|---|
| QuickGO |
En biomembran er et separasjonslag som omgir et cellerom eller, som en cellemembran, skiller det indre av en celle fra utsiden. Innenfor cellen skiller ulikt strukturerte biomembraner det indre av organeller eller vakuoler fra cytoplasma . En biomembran har en aktiv rolle gjennom membrankomponenter i den selektive transporten av molekyler og overføring av informasjon mellom de to avdelingene som denne biomembranen befinner seg mellom.
permeabilitet
Siden biomembranen først og fremst er et separasjonslag mellom forskjellige områder, er den ugjennomtrengelig for de fleste molekyler. Mindre lipofile molekyler som karbondioksid , alkoholer og urea kan fritt diffundere gjennom lipidbilaget i membranen. For å tillate membranens permeabilitet for lipofobe partikler som vann eller store partikler som ioner eller sukkermolekyler , er forskjellige transportproteiner innebygd i membranen, som er ansvarlig for transport av visse stoffer. Det er derfor man snakker om selektiv permeabilitet [1] .
konstruksjon
En biomembran er alltid topologisk lukket og omslutter et mellomrom. Ikke-frittstående membraner forekommer ikke i intakte celler. Biomembraner er asymmetriske: de har en plasmatisk side som vender mot cytoplasma (P-side) og en ekstraplasmisk side (E-side). [2]
Biomembraner består av lipider og proteiner . Karbohydratkjeder kan knyttes til proteinene. Som et lipiddobbelt lag danner lipidkomponenten membrans grunnstoff og er ansvarlig for dets spesielle fysisk -kjemiske egenskaper. Spesielt fungerer dette dobbeltlaget som et passivt separasjonslag. Steroider som kolesterol inngår i en hydrofob interaksjon med lipidene og størkner den ellers fleksible biomembranen ved høye konsentrasjoner i biomembranen. I tillegg distribueres proteiner på og i membranen, som tar på seg de aktive funksjonene til membranen. Proteinene har bare en veldig liten støttefunksjon for biomembranen, da de svømmer gjennom lipidlagene.
Biomembraner kan karakteriseres basert på dens tetthet ; den er vanligvis mellom 1,12 og 1,22 g · cm −3 . Tettheten avhenger av vektforholdet mellom proteinene og lipidene: avhengig av membranets funksjon, verdier på 0,25 ( myelinmembran , lavt proteininnhold), 1,3 (plasmamembran av erytrocytter ), 2,5 (plasmamembran av E . coli ), 2,9 (indre mitokondriell membran ) opp til en verdi på 5 i den lilla membranen som finnes i Halobacterium (høyt proteininnhold). [3]
I visse typer celleorganeller ( kjerne , mitokondrion , plastid ) vises biomembraner som doble membraner .
Fosfolipid dobbeltlag
Lipiddobbeltlaget består stort sett av amfifile fosfolipider , som har en hydrofil hodegruppe og en hydrofob halegruppe (for det meste hydrokarbonkjeder ). Som et resultat av den hydrofobe effekten dannes et dobbeltlag i vann, med de hydrofobe halene pekende innover og de hydrofile hodene pekende utover. På grunn av den hydrofobe kjernen er et slikt lipiddobbelt lag nesten ugjennomtrengelig for vann og vannløselige molekyler, men samtidig veldig fleksibelt og mekanisk vanskelig å ødelegge. Av denne grunn forlater ikke engang en punktering med en pipette et hull i membranen. For å gjøre dette kan det bli ødelagt av lipidløsningsmidler og lipaser .
Membraner består av tre hovedtyper av lipider: fosfoglyserider, sfingolipider og kolesterol.
Fosfolipider
- Fosfolipider er preget av en fosfatgruppe , de utgjør hoveddelen av membranlipider. For det meste har de en grunnleggende struktur laget av glyserin , "over" membranen, og derfor kalles de fosfoglyserider. To av de tre hydroksylgruppene av glyserol er forestret med hydrofobe fettsyrer, den tredje med en hydrofil fosfatgruppe. Fosfatgruppen kan bære en ytterligere substituent. Hvis den ikke gjør det som nesten aldri forekommer i membraner, vil molekylet bli kalt fosfatidsyre . En hyppig substituent er kolin , som fører til fosfatidylkolin (PC), eller etanolamin , som fører til fosfatidyletanolamin (PE), serin, fører til fosfatidylserin (PS) eller inositol , fører til fosfatidylinositol (PI). Det gjelder at alle de beskrevne molekylene består av en hydrofil hodegruppe, fosfatet med substituenter og en hydrofob hale, en uforgrenet fettsyre med 16 til 20 atomer. Avhengig av antall dobbeltbindinger i fettsyren, skilles det mellom mettede fettsyrer (ingen dobbeltbindinger ), enumettede (en dobbeltbinding) og flerumettede.
Shingolipider
- En sfingolipid er en forbindelse av en sfingosin som er knyttet til en fettsyre via aminogruppen. Hydroksylgruppen kan esterifiseres med forskjellige grupper, uten forestring av ceramider , esterifisering med fosfokolin gir sfingomyelin og med sakkarider glykosofingolipider . Shingolipider er også amfipatiske og ligner fosfolipider i denne forbindelse.
kolesterol
- Dyremembraner kan inneholde opptil 50% kolesterol ( vektprosent ), mindre i planter og ikke i det hele tatt i bakterier. Kolesterol er lite og ikke særlig amfipatisk, og det er derfor bare hydroksylgruppen er på membranoverflaten og resten av molekylet er i membranen. Det stive ringsystemet av kolesterol hindrer strømmen av lipidlaget, noe som gjør det mer stivt. [4]

Lipid -dobbeltlaget i en biomembran er normalt flytende, noe som betyr at lipidene og proteinene er ganske mobile i membranplanet. Imidlertid er en utveksling av lipider mellom de to lagene eller til og med en løsrivelse av et lipid fra membranen svært sjelden. En målrettet bevegelse fra den ene siden av membranen til den andre ( flip-flop ) er normalt bare mulig med aktiv deltakelse av spesielle proteiner (såkalte flippaser og floppaser ) mens du bruker adenosintrifosfat (ATP). Flippaser transporterer lipider fra utsiden av plasmamembranen til den cytosoliske siden. Floppaser er klassiske ABC -transportører og transporterer membranlipider fra den cytosoliske siden av plasmamembranen til utsiden. Ytterligere transportører for membranlipider er scram bobler , som imidlertid ikke utveksler membranlipider i retning av konsentrasjonsgradienten på en ATP-avhengig måte før en likevekt er etablert.
Hvor flytende lipiddobbeltlaget er, avhenger først og fremst av antall dobbeltbindinger i de hydrofobe hydrokarbonkjedene til lipidene; noen bakterier [5] bruker også forgrenede kjeder. Jo mer, jo mer væske membranen er. På den annen side bestemmes væskegraden av andre innebygde molekyler. Kolesterol reduserer for eksempel på den ene siden fluiditet, men ved lave temperaturer forhindrer det at membranen størkner som en gel .
Vitamin E er en antioksidant (som vitamin C ); det beskytter de umettede hydrokarbonkjedene til fosfolipidene i biomembranen fra å bli ødelagt av frie radikaler ( lipidperoksydasjon ).
Membranproteiner

Ulike typer membranproteiner som er innebygd i lipiddobbeltlaget sikrer forskjellige egenskaper til biomembranen gjennom protein-lipid-interaksjoner . De to sidene av en biomembran kan også variere sterkt på grunn av arrangementet av membranproteinene. For eksempel, mens reseptorer for celle-cellekommunikasjon og for påvisning av miljøendringer er rettet utover, peker enzymer som er involvert i reaksjoner innover (dvs. de er lokalisert i cytoplasma).
Mange proteiner er involvert i membrantransport , dvs. i utveksling av stoffer og signaloverføring via spesifikke reseptorer. Et stort antall membranproteiner som kjennetegner forskjellige celletyper og modenhetstrinn og som kan variere fra individ til individ ( f.eks. Blod- og vevsgrupper ) har blitt godt studert. Dette inkluderer også molekyler (for det meste glykoproteiner) som bidrar til skillet mellom seg selv og andre i henhold til lås -og -nøkkel -prinsippet .
I følge den flytende mosaikkmodellen er membranproteinene ikke stivt festet i membranen, men er i stand til svært dynamiske endringer i plasseringen i membranen. Denne dynamikken danner forutsetningen for utløsning av signalkjeder på mobilnivå, både intracellulært og mellom samarbeidende celler.
Membranproteinene kan klassifiseres i henhold til deres forankring i lipiddobbeltlaget:
Integrerte proteiner
- Gensekvensering antyder at 30% av alle kodede proteiner er integrerte proteiner. Som transmembrane proteiner krysser integrerte proteiner begge lagene i lipidbilaget, noen ganske enkelt, andre med flere sløyfer. Deler av proteinet stikker ut fra membranen. Integrerte proteiner, som fosfolipider, er amfipatiske. Domener i membranen er hydrofobe, aminosyreresten interagerer med lipidkjedene. Imidlertid ville disse ustyrte kreftene alene ikke være tilstrekkelige for stabilisering; I mange proteiner samhandler en stripe med stort sett basiske rester med de ladede hodegruppene til fosfolipidene. Den andre delen som stikker ut fra membranen samhandler med det omkringliggende vannet og stoffene som er oppløst i det. Integrerte proteiner er ikke nødvendigvis godt forankret i membranen, men kan også være fritt mobile.
Perifere proteiner
- Perifere proteiner finnes på innsiden og utsiden av membranen [6] . De er midlertidig knyttet til disse eller til integrerte proteiner gjennom en blanding av elektrostatiske og hydrofobe interaksjoner, så vel som andre, ikke-kovalente bindinger. Vedlegget er dynamisk, avhengig av tilstanden, kan det bindes eller løsnes. Membranen trenger ikke å bli ødelagt for å få den; en sterkt konsentrert saltoppløsning er tilstrekkelig til å bringe dem i løsning, da dette svekker de elektrostatiske interaksjonene. Som et eksempel er det best studerte på den cytoplasmatiske siden proteiner som, som fibriller, danner noe som et skjelett, de som danner belegg og enzymer. Perifere proteiner utenfor tilhører for det meste den ekstracellulære matrisen. Integrale og perifere proteiner kan modifiseres post-translasjonelt ved binding til fettsyrerester, prenylering eller et GPI-anker .
Lipidforankrede proteiner
- Lipidforankrede proteiner er perifere proteiner og stikker derfor heller ikke ut gjennom membranen, men er kovalent knyttet til et lipidmolekyl som er innebygd i membranen. Det skilles mellom forskjellige typer (inkludert prenylering ( farnesylering , geranylgeranylering ), S-acylering eller myristoylering), men mange er GPI-forankret . Proteiner med et GPI -anker er plassert på utsiden av plasmamembranen. [4]
funksjon
Cytoplasma inne i en celle skilles fra utsiden av en biomembran. Disse kalles cellemembraner , plasmamembraner , plasmalemmer eller membrana cellularis . Biomembraner har følgende oppgaver:
- Kompartementalisering
- Av energiske årsaker representerer hver biomembran et gapfritt lag.Med flere membraner er det automatisk separate mellomrom, såkalte rom. De fleste celler inneholder reaksjons- og lagringsrom ( rom ), for eksempel celleorganeller og vakuoler med svært forskjellige kjemiske egenskaper. Det er veldig forskjellige stoffer i de forskjellige rommene. Således veldig forskjellig, z. Noen ganger er til og med motsatte prosesser mulig på samme tid som ikke påvirker hverandre, for eksempel oppbygging og nedbrytning av karbohydrater. Videre er individuell regulering mulig.
- Stillas for biokjemisk aktivitet
- For spesifikke reaksjoner er den nøyaktige justeringen av molekylene i forhold til hverandre nødvendig, da visse interaksjoner må inngås. Denne nøyaktige justeringen er ikke mulig i løsning. Biomembraner gir nå et rammeverk for hvilke molekyler effektivt kan samhandle og reagere med hverandre. Ellers ville ikke viktige reaksjoner vært mulige; multi-enzymkomplekset i respirasjonskjeden og fotosyntesen , for eksempel, er forankret i membranen.
- Selektiv permeabilitet
- Partikler trenger ikke hindre membraner uhindret, men kan velges og eventuelt beholdes.
- Transport av oppløste stoffer
- Molekyler kan transporteres fra den ene siden av membranen til den andre, selv mot en konsentrasjonsgradient (dvs. aktivt). På denne måten kan næringsstoffer bli beriket i cellen. Ioner kan også transporteres over membranen, som spiller en stor rolle i nerver og muskler.
- Reaksjon på eksterne signaler
- Plasmamembranen er viktig for en reaksjon på ytre stimuli (dvs. for signaloverføring). Det er reseptorer i membranen. Hvis et bestemt molekyl diffunderer i nærheten (en ligand), kan begge kombineres på grunn av deres komplementære struktur og reseptoren sender et signal til cellen. Ulike reseptorer gjenkjenner forskjellige ligander slik at cellen kan absorbere informasjon om omgivelsene. Reaksjoner på miljøet må tilpasse metabolismen ved å endre enzymaktiviteten, frigjøre lagringsmaterialer eller til og med begå selvmord.
- Intercellulær interaksjon
- Plasmamembranen er det ytre laget av cellen. I flercellede celler samhandler en celle med sine naboceller via plasmamembranen. Så celler kan f.eks. B. holde sammen eller utveksle signaler og materiale.
- Energiomstilling
- Membraner er involvert i energikonverteringer som fotosyntese og nedbrytning av karbohydrater. I eukaryoter foregår førstnevnte i kloroplastene , sistnevnte i mitokondriene .
- Forstørrelse av overflaten
- Små fremspring av biomembranen, såkalte microvilli , forstørrer celleoverflaten og dermed området som kan arbeides med, ettersom metabolismen skjer spesielt intensivt på biomembranen. [4]
Flytende
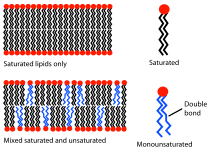
Fluiditeten til en biomembran avhenger av temperaturen . En membran laget av fosfatidylkolin og fosfatidyletanolamin, hvis fettsyrerester er mettet, vil være ganske flytende ved 37 ° C. I denne tilstanden kan membranen sees på som et todimensjonalt flytende krystall . De lange aksene til fosfolipidene er parallelle på linje, fosfolipidene selv kan rotere og bevege seg fritt i planet. Opp til en viss temperatur, overgangstemperaturen, bevegelsen av fosfolipidene er sterkt begrenset og membranegenskapene endres, tilstanden ligner nå den på en frossen gel . Overgangstemperaturen avhenger av typen lipider; jo kortere de er og jo flere dobbeltbindinger de inneholder, jo lavere er den. Kolesterol forstyrrer membranens normale struktur og reduserer mobiliteten til membranlipider. Overgangstemperaturen kan da ikke lenger bestemmes klart. I dyreceller sikrer lipidkolesterolet at væskenivået opprettholdes, selv med temperatursvingninger.
betydning
Fluiditeten til en biomembran ligger mellom stiv og flytende og tillater dermed en viss struktur. Membranproteiner kan samles i funksjonelle enheter og senere skilles igjen. Dette er for eksempel viktig for fotosyntese. Fluiditet spiller også en stor rolle i membrangenerering og er viktig for mange grunnleggende prosesser som celledeling , cellevekst , sekresjon, etc. Mens temperaturen ofte svinger, må membranens fluiditet forbli konstant. For å oppnå dette kan membranlipidene modifiseres: en utveksling av fosfolipider er mulig; Desaturaser kan danne dobbeltbindinger fra enkeltbindinger, fosfatryggraden og lipidhalene til fosfolipidene kan omfordeles og en høyere andel umettede fettsyrer kan produseres enn før. På denne måten kan spesielt kaldblodige skapninger tilpasse seg miljøet.
Lipidflåter
I biomembranen er ikke lipidmolekyler jevnt fordelt, men det er mikrodomener med en spesiell lipidsammensetning. Kolesterol og sfingolipider er spesielt utsatt for en slik tilknytning. Noen proteiner, for eksempel de med GPI -ankre, akkumuleres i slike områder, mens andre er spesielt sjelden funnet der. Lipidflåter er trolig veldig små og i en konstant prosess med oppløsning og regenerering.
historie


- 1895 Charles Ernest Overton antar at biomembranene består av lipider. Dette konkluderte han med observasjoner om at lipofile (fettløselige) stoffer, for eksempel visse bedøvelsesmidler, lettere kan komme inn i cellene enn stoffer som er lipofobe.
- 1917 Irving Langmuir mistenker at fosfolipider flyter på overflaten av vannet.
- I 1925 utviklet de nederlandske forskerne Gorter og Grendel bilagsmodellen : [7] Fosfolipider med hydrofile grupper er arrangert som et dobbelt lag i membranen på en slik måte at de hydrofile gruppene i lipidene peker utover og de hydrofobe i det indre av dobbeltlaget. Men med deres modell så de to forskerne helt bort fra det store proteininnholdet i biomembranen.
- I 1935 presenterte JF Danielli og H. Davson den klassiske modellen for strukturen til en biomembran: Biomembranen består av et bimolekylært lipidlag . De hydrofobe halene til lipidene vender mot hverandre, de hydrofile hodene er belagt med proteiner . Kort sagt: protein - lipid dobbeltlag - protein (sandwichstruktur). Elektronmikroskopiske bilder av biomembraner avslører en trelags struktur: to ytre lag (hvert 2,5 nm tykt) og et mellomlag (3 nm tykt). Denne membranmodellen kalles enhetsmembran (ger.: Enhetsmembran).
- 1972 utviklet Seymour Jonathan Singer og Nicolson GL, fluidmosaikkmodellen (fluidmosaikkmodell) av en biomembran: [8] Globulære proteinmolekyler for å "flyte" i en bimolekylær lipidfilm. Lipidfilmen oppfører seg som en viskøs todimensjonal væske , så lipidmolekyler og proteiner kan diffundere uhindret i membranplanet. Det er to typer membranforening av proteiner. Integrerte proteiner, også kalt transmembrane proteiner, strekker seg gjennom membranen. Perifere proteiner , også kalt assosierte proteiner, blir avsatt på lipidbilaget.
- 1972: På samme tid som Singer og Nicolson konkluderte Frye og Edidin med forsøk med to celler der visse membranproteiner ble merket at membranen ikke kan være statisk, men er i konstant bevegelse. De kombinerte de merkede cellene og de markerte områdene av membranen som bare var tilstede separat blandet.
- I 1983 presenterte Mouritsen og Bloom madrassmodellen til cellemembranen. Den sier at den hydrofobe delen av membranproteinene som er innebygd i membranen ikke alltid er nøyaktig den samme størrelsen som cellemembranen, og at lipider med forskjellige kjedelengder derfor lagres passende rundt visse membranproteiner. [9]
- Siden etableringen av den flytende mosaikkmodellen av Singer og Nicholson i 1972, har det blitt oppdaget mange indikasjoner som førte til formuleringen av den dynamisk strukturerte mosaikkmodellen [10] . Ulike studier har vist at proteiner og forskjellige lipidmolekyler på ingen måte er jevnt fordelt på overflaten av membranen, som man kunne forvente i en ren væske. I stedet ser det ut til å være områder med høy konsentrasjon av visse proteiner (såkalte reseptorøyer ) eller visse typer lipider (såkalte lipidflåter ) som stadig omgrupperes, oppløses og kommer sammen igjen.
Indre membransystem
Det indre membransystemet består av forskjellige celledeler eller organeller i eukaryote celler, som er omgitt av biomembraner. Disse inkluderer: kjernemembranen , det endoplasmatiske retikulum , Golgi -apparatet , lysosomer , vesikler , endosomer og plasmamembranen . De fleste proteiner som er beregnet på organeller i det indre membransystemet, transporteres cotranslasjonelt inn i det endoplasmatiske retikulum og derfra via sekretorisk vei i det indre membransystemet.
weblenker
- Biomembraner / plasmamembran og glycocalyx / flytende mosaikk modellgrafikk
- Phillip Eichman: Fra lipidbilaget til væskemosaikken: en kort historie om membranmodeller. SHiPS Ressurssenter: Sosiologi, historie og vitenskapsfilosofi: lærernes nettverksnyheter. Bind 9 (2), 1999.
Individuelle bevis
- ^ Biomembrane I: Selektiv permeabilitet av membraner . ( Memento fra 8. februar 2008 i Internettarkivet )
- ↑ H. Kleinig, P. Sitte: Cell Biology. 2. utgave. Stuttgart 1986, ISBN 3-437-30528-X , s.33 .
- ↑ Hans Kleinig, Uwe Maier: Kleinig / tilpasset cellebiologi. Verlag Gustav Fischer, 1999, ISBN 3-437-26010-3 .
- ↑ a b c Gerald Karp, Kurt Begin: Molecular Cell Biology. Springer, 2005, ISBN 3-540-27466-9 , s. 157-230.
- ↑ T. Kaneda: Iso- og anteiso-fettsyrer i bakterier: biosyntese, funksjon og taksonomisk betydning. I: Mikrobiol. Rev. 55 (2), juni 1991, s. 288-302. PMID 1886522
- ^ S. Tan, HT Tan, MC Chung: Membranproteiner og membranproteomikk . I: Proteomics . teip 8 , nei. 19. oktober 2008, s. 3924-3932 , doi : 10.1002 / pmic.200800597 , PMID 18763712 .
- ^ E. Gorter, F. Grendel: På bimolekylære lag av lipoider på blodets kromocytter. I: Journal of Experimental Medicine . Bind 41, 1925, s. 439-443.
- ^ SJ Singer, GL Nicolson: Den flytende mosaikkmodellen for strukturen i cellemembraner. I: Vitenskap . Bind 175, 1972, s. 720-731. PMID 4333397 .
- ^ OG Mouritsen, M. Bloom: Madrassmodell av lipid-protein-interaksjoner i membraner . I: Biophys. J. Band 46 , nei. 2 , august 1984, s. 141–153 , doi : 10.1016 / S0006-3495 (84) 84007-2 , PMID 6478029 , PMC 1435039 (fri fulltekst).
- ↑ G. Vereb et al.: Dynamisk, men likevel strukturert: Cellemembranen tre tiår etter Singer-Nicolson-modellen. I: Proc. Natl. Acad. Sci. USA. Bind 100, 2003, s. 8053-8058. PMID 12832616 ,PMC 166180 (gratis fulltekst).
- Cellekomponent
- Cellebiologi