Cellemembran
| Forelder |
| celle |
| Genontologi |
|---|
| QuickGO |
Cellemembran ( anatom. Membrana cellularis ), cytomembran , plasmamembran , også kalt plasmalemma , er navnet på hver levende celles eget biomembran , som omgir celleinnredningen og opprettholder sitt indre miljø.

Cellemembranen til eukaryote celler og bakterier består av et dobbeltlag av lipider ( lipiddobbelt lag ) og forskjellige membranproteiner som er innebygd i det perifert, integrert eller transmembrant. De hydrofile , vannløselige polene i lipiddobbeltlaget danner den indre og ytre overflaten av cellemembranen, mens de hydrofobe , vannavvisende polene i lipiddobbeltlaget er rettet mot midten av membranen. Avhengig av størrelsen, fettløseligheten og ladningen av visse stoffer, er cellemembranen permeabel i forskjellige grader eller dårlig, så den er semipermeabel. [1] Med en tykkelse på omtrent seks til ti nanometer kan den bare vagt forestilles som en linje under et lysmikroskop . Cellemembranen skiller det cytoplasmatiske rommet til en celle fra det ekstracellulære rommet og representerer grensesnittet gjennom hvilket en utveksling av stoffer med miljøet finner sted.
I tillegg påtar cellemembranen seg viktige oppgaver i celle-cellekontakter , celle-matrise-tilkoblinger , signaloverføring , cellemigrasjon og endringer i celleform . I prokaryoter er det også stedet for fotosyntese eller den kjemotrofiske tilførselen av energi og arrangementsoverflate for fotosystemer , respirasjonskjedekomplekser og andre enzymsystemer . Den ytre, ekstracellulære siden av cellemembranen er også strukturelt forskjellig i alle cellene fra den indre, intracellulære siden av membranen. På utsiden kan plasmamembranen være omgitt av en cellevegg som et beskyttende deksel. Innad kan den kobles til proteiner fra et cytoskjelett, eller i noen encellede organismer kan den stivnes for å danne en pellikkel .
Organisering av cellemembranen
sammensetning
Cellemembranen består i utgangspunktet av et stort antall forskjellige membranogene lipider i to motstående lag, som sammen danner et lipiddobbelt lag; bare i noen archaea dannes det som et lipidmonosjikt . [2] Hovedkomponentene er fosfolipider , for det meste forskjellige fosfoglyserider - disse inkluderer lecitiner som fosfatidylkoliner (PC) og cefaliner som fosfatidyletanolaminer (PE) eller som fosfatidylseriner (PS) samt fosfatidylinositoler (PI). I tillegg til sphingomyelin , er andre viktige membrankomponenter også andre sfingolipider og også steroler som kolesterol . De ikke- steroide lipidkomponentene er forskjellige, bortsett fra hodegruppen, for eksempel i metningsgraden og kjedelengden av bundne fettsyrerester . I tillegg kan de ha blitt modifisert med karbohydratrester for å danne glykolipider .
I tillegg til lipider inneholder cellemembranen også en rekke spesifikke membranproteiner . De ligger på utsiden eller innsiden, og de har forskjellige oppgaver. Som et transmembranprotein strekker noen seg gjennom membranen og forbinder begge sider, for eksempel celleadhesjonsmolekyler , ionekanaler eller reseptorer for signaltransduksjon gjennom membranen. Måten de er utstyrt med membranproteiner karakteriserer en celle i henhold til visse funksjoner. Men selve sammensetningen av lipidene skiller cellemembranen fra andre cellemembraner som omslutter celleorganeller .
Som det ytterste laget på den menneskelige celleoverflaten danner antenneformede sukkerkjeder av glykoproteiner og glykolipider i cellemembranen-og muligens de i den ekstracellulære matrisen-et karbohydratrikt skall, det såkalte glykokaloksiet . Som den første barrieren beskytter dette cellen mot mekaniske og kjemiske effekter, formidler celle-celle-interaksjoner og forhindrer uønskede protein-protein-kontakter med andre celler. [3] [1]
polaritet
Lipidsammensetningen er forskjellig med hensyn til den intracellulære / cytosoliske siden og den ekstracellulære siden av cytomembranen. Denne orienterte organisasjonen er forårsaket av selektive fosfolipid -translokatorer , kalt flippaser eller floppaser . I løpet av apoptose reverseres denne orienteringen og fungerer som et signal for cellene rundt.
Lipidflåter
Sfingolipider og kolesterol kan danne domener i cellemembranen, de såkalte lipid flåter . Lipidflåter er selektivt beriket i visse membranproteiner i henhold til deres transmembrane domener og spiller en rolle i gruppering av membranproteiner, for eksempel i løpet av en signalkaskade . Det er foreløpig (2015) ingen konsensus om den typiske størrelsen og levetiden til lipidflåter.
eiendommer
Med cellemembranen avgrenser cellen sitt eget rom der andre miljøforhold er mulige enn i området rundt. I følge væskemosaikkmodellen ligger forskjellige membranproteiner som er viktige for eksterne kontakter i membranen eller "svømmer". Som reseptorproteiner på utsiden absorberer de stimuli eller mottar signaler; som membranspennende proteiner tillater de transport gjennom membranen eller forankrer cellen i miljøet eller kobler den til andre celler.
Mange av de utoverrettede perifere proteiner og noen av membranlipider ofte bære kortkjedede, noen ganger trelignende forgrenede karbohydratforbindelser , og er derfor også betegnet som glykoproteiner eller glykolipider . Totalt sett danner disse overflatestrukturene glykokalyksen , som sikrer stabilitet i tillegg til cytoskjelettet i celler uten cellevegg . En celle kan gjenkjennes av immunceller , for eksempel ved hjelp av karakteristiske strukturer, også kjent som overflateantigener - eller omvendt kan de identifisere og identifisere seg selv på denne måten.
Cellemembranen kan også binde molekyler, molekylære komplekser eller større partikler fra miljøet på en spesiell måte. I tillegg kan det oppstå deformasjoner av cellemembranen, slik som fordypninger. Hvis visse protein-lipid-interaksjoner resulterer i invagasjon og innsnevring av cellemembranseksjoner, er områder av det omkringliggende mediet innelukket og absorbert, som kalles endocytose . Her skilles det etter det vedlagte innholdet mellom fagocytose av faste stoffer eller andre organismer og pinocytose av væskemengder, eller etter volumet mellom matvakuoler og de mindre vesiklene .
Exocytose er navnet som gis til prosesser i motsatt retning, der membranen av vesikler smelter sammen med cytomembranen, og vesikelinnholdet blir dermed utvist ut i det omkringliggende ekstracellulære rommet. I eukaryote celler er dannelse av vesikler også mulig gjennom innsnevring av indre membraner, fra hulrom i det endoplasmatiske retikulum og Golgi -apparatet . Ved konstitutiv eksocytose brukes slike vesikler ikke til å skille ut spesifikke vesikkelinnhold, men for å utvide eller omforme cellemembranen og inkorporere membranproteiner.
Som endringer i cellens form kan områder av cellemembranen også stikke ut eller dessuten vende fremover som en celleprosess , støttet av et rammeverk av mikrotubuli i cytoskjelettet, for eksempel som cilia . I motsetning til disse er mikrovilliene spesielle formasjoner av dyrecellemembraner - i form av pseudopodia -lignende fremspring utover - som, båret av aktinfilamenter , forstørrer overflaten av membranen og sammen danner den såkalte penselgrensen til en celle. I encellede organismer som amøber er pseudopodene deres på lignende måte konstruert fremspringende celleprosesser som de beveger seg med.
Alle cellemembraner er halvgjennomtrengelige , de er gjennomtrengelige for noen stoffer og mindre eller slett ikke for andre. Selv om lipidlaget ikke representerer en barriere for lipofile molekyler, kan små hydrofile bare passere gjennom åpninger som porer eller kanaler - hvis de passer gjennom og ikke blir frastøtt som ladede ioner - og for større som glukose en støttet transport gjennom spesielle bærere er nødvendig for membranpassasjen. Bortsett fra i eksocytotiske vesikler, kan ikke proteinene i cytoplasma krysse cellemembranen. Det er derfor fordelingen av stoffer og ladninger inne i cellen er forskjellig fra utsiden. Dette skaper elektrokjemiske gradienter på tvers av membranen, noe som fører til en potensiell forskjell mellom innsiden og utsiden. Alle celler har derfor et membranpotensial , som kan justeres til et visst nivå, hovedsakelig ved bruk av ionekanaler og ionepumper .
Fra et elektrofysiologisk synspunkt kan cellemembranen ses i forenklet form som en platekondensator hvis spesifikke kapasitet for membraner av forskjellige celletyper er i området ca. 1 µ F · cm −2 . [4]
Kanaler, transportører, pumper og reseptorer
Cellemembranen har en hydrofil , vann-tiltrekkende, polar hodegruppe på ytre og indre overflate. Sentrum av cellemembranen kan forstås som hydrofob . Dette dobbeltlags lipidlaget er godt gjennomtrengelig for små molekyler som vann, og det er svært gjennomtrengelig for oppløste gasser ( O 2 , CO 2 , N 2 ). Siden membranen stort sett er ugjennomtrengelig for de fleste stoffer på grunn av molekylstørrelsen eller ladningen (ioner) [5] [1] [3] , kan det opprettes et spesielt miljø i cellen, som skiller seg fra det vandige, ytre miljøet i den ekstracellulære plassen er forskjellig. Stoffene som er oppløst i vannet kan passere gjennom membranen gjennom spesielle porer, kanaler eller transportører, hvis type og antall bestemmes av cellen. [6]
kanaler
Ionkanaler er proteiner som når gjennom cellemembranen og danner en hydrofil kanal som stort sett ioner kan passere gjennom. Transporten er passiv, ionene flyter enten inn i eller ut av cellen etter den elektrokjemiske gradienten . Denne prosessen skjer spontant (med en reduksjon i fri entalpi er den eksergonisk ). Åpningen og lukkingen av en kanal kan vanligvis kontrolleres og er z. B. styrt av elektrisk spenning eller signalstoffer. [7]
De fleste celler har også spesifikke kanaler for vann, såkalte aquaporiner . [7]
Transportør
Transportører eller bærere lar ioner eller små hydrofile molekyler som glukose eller aminosyrer passere. De føres gjennom membranen av de respektive transportproteinene som gjennomgår en konformasjonsendring , og veksler mellom konformasjoner hvis bindingssteder er på den ene eller den andre siden av membranen. Bare hvis det er en konsentrasjonsgradient for det respektive stoffet, skjer transporten på en balansert måte ( Uniport ), og det er derfor det her omtales som lettere diffusjon . [7]
Biltransportøren kan tilordnes transportøren eller ses på som uavhengig. I prinsippet er det en bærer som transporterer to stoffer gjennom membranen samtidig, enten i samme retning ( symport ) eller i motsatt retning ( antiport ). Det ene stoffet følger stort sett passivt en gradient, hvis drivkraft også kan brukes til å transportere det andre stoffet mot en gradient. I dette tilfellet snakker man også om sekundær aktiv transport . Eksempler er natrium-glukose-symporten i tarmepitelet eller natrium-hydrogen-antiporten i det proksimale tubuli . [Åttende]
pumpe
Pumper transporterer stoffer mot en gradient gjennom cellemembranen og bygger dermed opp forskjeller i distribusjon. Denne hovedsakelig aktive transporten koster hydrolysen av ATP . Derfor er disse pumpene også enzymer som deler ATP og bruker frigjort Gibbs -energi til transport, og blir derfor også referert til som transport -ATPase . Eksempler på dette er ionepumper som natriumkaliumpumpen eller kalsiumpumpen i det endoplasmatiske retikulum . [9]
Archaea cellemembran
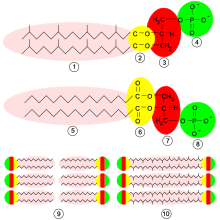
I archaea viser cellemembranen ofte en spesiell struktur. Lipidkomponentene er ofte ikke fettsyrer, men isoprenoidalkoholer , som ikke er knyttet til glyserol gjennom en ester, men snarere en eterdannelse . Det er også isoprenoider, som danner bindinger på begge sider og (som bololipider ) og dermed trekker gjennom og spenner membranen som forbinder begge lagene i lipidlaget. [10] Cellemembranen til disse organismene er ordnet i et enkelt lag med slike di- og tetraetere og er mye mer temperatur og syre-stabil. [11] Dette gjør at archaea kan kolonisere habitater med ekstreme miljøforhold, for eksempel varme termiske kilder .
weblenker
- Plasmamembran vist som en flytende mosaikkmodell med struktur og funksjon i forbindelse med cellebiologi - grafikk
- Thumbnail oversikt cellemembran (Membrana cellularis)
Individuelle bevis
- ↑ a b c Stephan Dönitz: Menneskekroppssykdom for redningstjenesten . Red .: Frank Flake. 1. utgave. Elsevier, 2015, ISBN 978-3-437-46201-6 , s. 37 .
- ^ Frank Hoffmann: Karakterisering av kirale Langmuir -filmer ved hjelp av IR -spektroskopiske, termodynamiske og avbildningsmetoder samt biomimetiske tilnærminger for å belyse virkningsmåten til antibiotika peptidet surfactin. Diss. Univ. Hamburg 2002, s. 6 ( d-nb.info PDF).
- ↑ a b Agamemnon Despopoulos: Pocket Atlas Physiology . 8., revidert. og eksp. Utgave. Thieme, Stuttgart 2012, ISBN 978-3-13-567708-8 .
- ↑ Gerold Adam, Peter Läuger, Günther Stark: Fysisk kjemi og biofysikk . 5. utgave. Springer-Verlag, 2009, ISBN 978-3-642-00424-7 , s. 359 ( begrenset forhåndsvisning i Google boksøk).
- ^ Günther Jung: Organisk kjemi. Grunnleggende, sammensatte klasser, reaksjoner, konsepter, molekylær struktur, naturlige stoffer . 6., revidert. Utgave. Thieme, Stuttgart 2009, ISBN 978-3-13-541506-2 , s. 936 (med 133 bord).
- ↑ Renate Lüllmann-Rauch i samarbeid med Esther Asan: Pocket textbook histology . 5. utgave. Thieme, Stuttgart 2015, ISBN 978-3-13-129245-2 , s. 11 .
- ↑ a b c Renate Lüllmann-Rauch i samarbeid med Esther Asan: Pocket textbook histology . 5. utgave. Thieme, Stuttgart 2015, ISBN 978-3-13-129245-2 , s. 12.
- ↑ Renate Lüllmann-Rauch i samarbeid med Esther Asan: Pocket textbook histology . 5. utgave. Thieme, Stuttgart 2015, ISBN 978-3-13-129245-2 , s. 12 og 13 .
- ↑ Renate Lüllmann-Rauch i samarbeid med Esther Asan: Pocket textbook histology . 5. utgave. Thime, Stuttgart 2015, ISBN 978-3-13-129245-2 , s. 13.
- ^ A. Pearson, Y. Pi, W. Zhao, W. Li, Y. Li, W. Inskeep, A. Perevalova, C. Romanek, S. Li, C. Zhang: Faktorer som styrer fordelingen av arkeiske tetraethers i terrestriske Varme kilder . I: Anvendt og miljømikrobiologi . teip 74 , nei. 11. juni 2008, ISSN 0099-2240 , s. 3523-3532 , doi : 10.1128 / AEM.02450-07 , PMID 18390673 ( asm.org [PDF]).
- ↑ E. Boyd, T. Hamilton, J. Wang, L. He, C. Zhang: Tetraether -lipidsammensetningens rolle i tilpasningen av termofile arkaer til surhet . I: Grenser i mikrobiologi . teip 4 , nei. 62 , april 2013, ISSN 1664-302X , doi : 10.3389 / fmicb.2013.00062 , PMID 23565112 , PMC 3615187 (gratis tekst).
- Cellekomponent