Ribonukleinsyre

Ribonukleinsyre ( ribonukleinsyre ; kort RNS ; engelsk RNA for ribonukleinsyre ) (lat.-fr.-gr. oppbygd ord ) er en nukleinsyre som består av en kjede av mange nukleotider som et polynukleotid . I visse virustyper ( RNA -virus , retrovirus ) og de hypotetiske opprinnelige ribocyttene er biomolekylet bæreren av den genetiske informasjonen , dvs. materialets grunnlag for genene . Ordet består av ribose og nukleinsyre .
En vesentlig funksjon av RNA i den biologiske cellen er konvertering av genetisk informasjon til proteiner (se proteinbiosyntese , transkripsjon og translasjon ), i form av mRNA fungerer den som en bærer av informasjon. I tillegg utfører spesielle typer RNA andre oppgaver; når det gjelder RNA -virus , utgjør det til og med selve genomet . Videre består deler av cellekomponentene som er ansvarlig for konverteringen av denne informasjonen også av RNA: snRNA og snoRNA er involvert i modning av mRNA, de katalytiske komponentene i ribosomene danner rRNA , og tRNA transporterer byggesteinene for proteiner. Videre er spesielle RNA -er involvert i genregulering .
RNA kan også overta oppgavene til enzymer ( ribozym ) eller handle på samme måte som antistoffer ( aptamer ).
Struktur og forskjell på DNA
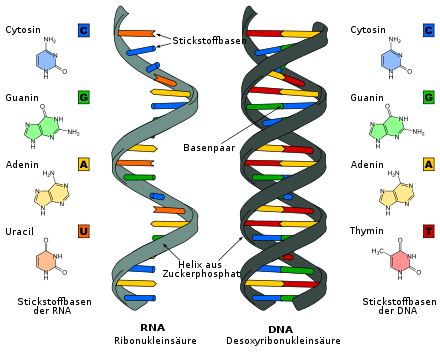
Når det gjelder struktur, ligner RNA på DNA . I motsetning til dobbeltstrenget DNA er RNA-molekyler vanligvis enkelttrådede, men kan danne karakteristiske refoldinger i korte strekninger med komplementære basesekvenser (AU, GC), som gir inntrykk av en dobbeltstrenget helix intramolekylært. Begge er polynukleotider der nukleobasene på sukker er knyttet til hverandre via fosforsyre -diestere. Enkeltstrengethet øker antall muligheter for tredimensjonale strukturer i RNA og lar den gjennomgå kjemiske reaksjoner som DNA ikke kan. I RNA består hvert nukleotid av en ribose (dvs. en pentose : et sukker med fem karbonatomer), en fosfatrest og en organisk base . Ribosen til RNA er identisk med DNA, bortsett fra en hydroksylgruppe i stedet for et hydrogenatom ved 2' -stillingen i pentosering (derav også deoksy ribonukleinsyre, D NA). Denne forskjellen gjør RNA mindre stabilt sammenlignet med DNA, ettersom det muliggjør hydrolyse av baser: OH -gruppen ved 2' -posisjonen til sukkeret blir fratatt protonen av et negativt ladet hydroksidion og oksygenet som blir etterlatt mister en Ringbinding med fosforet, hvorved bindingen til det neste nukleotidet brytes. RNA brytes ned i sine nukleotider.
Følgende organiske baser forekommer i RNA: adenin , guanin , cytosin og uracil . De tre første basene finnes også i DNA. Uracil, derimot, erstatter tymin som en komplementær base for adenin. RNA bruker antagelig uracil fordi det er mindre energisk å produsere (ingen metylsubstitusjon).
Hårnål, stamme og sløyfestrukturer er kjent som sekundære strukturer i RNA , men en spiralforming er også mulig, med hårnål- og stilkesløyfestrukturer som har både enkeltstrengede og dobbeltstrengede områder. Sløyfestrukturen angir enkeltstrengede sløyfestrukturer i et molekyl.
I likhet med DNA kan RNA også være tilstede som et dobbeltstrenget molekyl. Den har de typiske egenskapene til en Watson-Crick-helix: anti-parallelt arrangement av RNA-strengene og en høyrehendt helix. Den har form av en A eller A 'helix (se DNA ). A-RNA blir også referert til som RNA-11, homologt med A'-RNA, som omtales som RNA-12. Tallet etter bindestrek indikerer antall basepar per helixomdreining. A´-RNA forekommer ofte ved høye saltkonsentrasjoner (over 20%).
A-RNA: 11 basepar per helix-sving, pitch 2,7 nm til 2,8 nm, hellingsvinkel til spiralaksen ca. 14 °
A´-RNA: 12 basepar per helix-sving, stigning 3 nm, hellingsvinkel til spiralaksen 16 ° til 19 °
Enantiomeren av RNA som finnes i levende ting er D- RNA. Den består av D -ribonukleotider. Senterene for kiralitet er i D- ribose. Ved å bruke L -ribose kan L eller L -Ribonukleotiden til RNA syntetiseres. Dette er relativt mer stabilt sammenlignet med enzymatisk nedbrytning av RNases . [1]
Tertiær struktur
Nukleinsyrer kan også adoptere komplekse romlige strukturer: tRNA må være i riktig tertiær struktur for deres funksjon.
-

Tertiær og sekundær struktur (nederst til høyre på bildet) av et tRNA
-

Tertiær struktur av en pseudo-node


Syntese av RNA
Enzymet RNA -polymerase katalyserer DNA gjennom transkripsjonsprosessen fra nukleosidtrifosfat (NTP) RNA. For dette fester RNA -polymerasen seg til en nukleotidsekvens av DNAet kalt en promotor ( transkripsjonsinitiering ). Deretter skiller den DNA dobbeltspiralen i to enkelt DNA -tråder ved å bryte hydrogenbindinger i et kort område. Komplementære ribonukleotider er festet til den kodogene DNA -strengen gjennom baseparring . Ved eliminering av et pyrofosfat er de knyttet til hverandre ved en esterlignende binding mellom fosforsyre og ribose . Leseretningen til DNA går fra 3' -enden til 5' -enden, syntesen av det komplementære RNA tilsvarende 5 '→ 3'. Åpningen av DNA -dobbeltspiralen skjer bare i et kort område, slik at den delen av RNA som allerede er syntetisert henger ut av denne åpningen, med 5' -enden av RNA først. Syntesen av RNA avsluttes ved et DNA -segment som kalles en terminator. RNA -transkriptet frigjøres deretter og RNA -polymerasen løsner fra DNA.
RNA kan produseres kunstig ved fosforamiditt -syntese .
Biologisk betydning
RNA -molekyler kan utføre forskjellige funksjoner. RNA kan overføre genetisk informasjon. Andre RNA -molekyler hjelper til med å oversette denne informasjonen til proteiner og regulere gener . I tillegg kan RNA også ha katalytiske funksjoner som ligner på et enzym . RNA får derfor forskjellige navn, avhengig av funksjonen. Forrige små bokstaver indikerer de forskjellige typene RNA: [2]
- MRNA , messenger RNA (engl. Messenger RNA) kopierer informasjonen i et gen som ligger på DNA og overfører den til ribosomet , der ved hjelp av denne informasjonen kan proteinsyntese finne sted. I hvert tilfelle danner tre nukleotider som ligger ved siden av hverandre i leserammen til polynukleotidstrengen et kodon ved hjelp av hvilken en bestemt aminosyre som skal inkorporeres i et protein kan bestemmes klart. Denne forbindelsen ble funnet i 1961 av Heinrich Matthaei og Marshall Warren Nirenberg . Avkodingen av den genetiske koden markerer en ny begynnelse i nesten alle biovitenskap.
- Nukleosid-modifisert mRNA er en syntetisk, kjemisk modifisert messenger-ribonukleinsyre (mRNA) der individuelle nukleosider erstattes av andre naturlig modifiserte nukleosider eller av syntetiske nukleosidanaloger. Det brukes eksperimentelt eller terapeutisk.
Følgende klasser av RNA blir ofte referert til som ikke-kodende ribonukleinsyrer .
- AsRNA , antisense RNA , brukes til å regulere genuttrykk .
- CircRNA , sirkulært RNA, er involvert i reguleringen ved å binde seg til miRNA. [3]
- HnRNA , heterogent kjernefysisk RNA, forekommer i cellekjernen til eukaryoter og er en forløper for det modne mRNA, og derfor blir det ofte referert til som pre-mRNA (eller pre-mRNA for forløper-mRNA).
- MiRNAene , mikroRNAene er nært beslektet med siRNAene og brukes til å regulere slike prosesser. B. Spredning og celledød.
- Ribosbryterne brukes til genregulering . De kan enten ha en aktiverende eller undertrykkende effekt.
- Ribozymene er katalytisk aktive RNA -molekyler. Som enzymer katalyserer de kjemiske reaksjoner.
- RRNA , ribosomalt RNA , i likhet med tRNA, bærer ingen genetisk informasjon, men er involvert i konstruksjonen av ribosomet og er også katalytisk aktiv i dannelsen av peptidbindingen .
- SaRNA , selvforsterkende RNA , brukes i RNA-vaksiner for å forlenge virkningstiden.
- SiRNA , lite interfererende RNA , stammer fra en signalbane til cellen, som er oppsummert som RNAi (RNA Interference). Her brytes dsRNA (dobbeltstrenget RNA) opp av enzymet Dicer i mange mindre fragmenter på ca. 22 nukleotider i lengde ( siRNAene ) og inkorporeres i enzymkomplekset RISC (RNA-indusert lyddempingskompleks). Ved hjelp av de inkorporerte RNA -fragmentene binder RISC komplementært til DNA, f.eks. B. genområder, eller mRNA og kan "slå av" dem. siRNA -er forskes for tiden (2006) intensivt for sitt engasjement i forskjellige celleprosesser og sykdommer.
- ShRNA brukes til RNAi.
- SnoRNA , lite nukleolært RNA , finnes i nucleolus og de nært beslektede scaRNAene i Cajal -kroppene .
- SnRNA , lite kjernefysisk RNA , i kjernen av eukaryoter er ansvarlig for å spleise hnRNA ved spliceosomet .
- LncRNA , langt ikke-kodende RNA , er lengre enn 200 nukleotider og skiller seg dermed fra små regulatoriske RNA som miRNA og siRNA. [4]
- PiRNA , Piwi-interagerende RNA , er 26–31 nukleotider lange og skiller seg dermed fra de noe mindre miRNAene og siRNA-ene. De danner komplekser med PIWI-proteiner som er involvert i epigenetisk og post-transkripsjonell demping i kimceller. [5]
- TRNA , overførings -RNA , koder ikke for noen genetisk informasjon, men fungerer som et hjelpemolekyl i proteinsyntese ved å ta opp en enkelt aminosyre fra cytoplasma og transportere den til ribosomet. TRNA er kodet av et spesifikt RNA -gen .
- TracrRNA , som spiller en viktig rolle i CRISPR / Cas9 -systemet.
I flertallet av levende vesener spiller RNA en underordnet rolle for DNA som informasjonsbærer: Her er DNA det permanente lagringsmediet for genetisk informasjon, mens RNA fungerer som en midlertidig butikk. Bare RNA -virus (flertallet av alle virus) bruker RNA i stedet for DNA som permanent lagringsmedium. Det skilles mellom følgende typer RNA for taksonomi av virus :
* dsRNA : dobbeltstrenget RNA;
* ss (+) RNA : enkeltstrenget RNA brukt som mRNA;
* ss (-) RNA : enkeltstrenget RNA som fungerer som en mal for mRNA-produksjon.
I tillegg bruker noen virus RNA som et replikasjonsmellomprodukt (f.eks. Hepadnavirus ).
Nedbrytning av RNA
Siden det stadig dannes nytt RNA og siden forskjellige transkripsjoner kreves på forskjellige tidspunkter (differensielt genuttrykk), må ikke RNA i cellen være for stabilt, men må også utsettes for nedbrytning. Dette gjøres ved hjelp av RNases , enzymer som skiller forbindelsene mellom sukkerstrukturen til RNA og dermed danner monomerer (eller oligomerer), som kan brukes igjen for å danne nytt RNA. Når et RNA skal nedbrytes bestemmes hovedsakelig (men ikke utelukkende) av lengden på poly-A halen , som gradvis forkortes etter hvert som RNA forblir i cytoplasma. Hvis lengden på denne halen faller under en kritisk verdi, blir RNA raskt degradert. I tillegg kan RNA inneholde stabiliserende eller destabiliserende elementer som muliggjør ytterligere regulering.
I hvert fall når det gjelder mRNA fra eukaryoter , finner ikke RNA-nedbrytningen sted et sted i cytoplasma , men i de såkalte "P-Bodies" ( behandlingsorganer ), som er veldig rike på RNaser og andre som er involvert i RNA -omsetning (nedbrytning) Enzymer er. Sammen med Stress Granules fortsetter disse kroppene å tjene til kortsiktig lagring av mRNA og dermed igjen demonstrere den nære koblingen mellom RNA-metabolisme (her oversettelse og RNA-nedbrytning).
RNA -verdenshypotesen
RNA -verdenshypotesen sier at RNA -molekyler ble antatt å ha vært forløperne til organismer i kjemisk evolusjon . Hypotesen kan stammer fra RNAs evne til å lagre, overføre og reprodusere genetisk informasjon, så vel som fra dets evne til å katalysere reaksjoner som ribozymer . I et evolusjonært miljø ville disse RNA -molekylene forekomme oftere som fortrinnsvis reproduserer seg selv.
Nobelpriser
Flere nobelpriser har allerede blitt tildelt for forskning på RNA:
- I 1959 mottok S. Ochoa og A. Kornberg Nobelprisen i medisin for sine studier om syntese av RNA ved RNA -polymeraser .
- For deres oppdagelse av den katalytiske aktiviteten til RNA -molekyler (jf. Ribozym ), ble S. Altman og T. Cech hedret med Nobelprisen i kjemi i 1989.
- I 1993 mottok R. Roberts og P. Sharp Nobelprisen i medisin for sine studier om behandling av RNA i eukaryoter (se også spleising (biologi) ).
- I 2006 mottok Andrew Fire og Craig Mello Nobelprisen i medisin for oppdagelsen av RNA -interferens ; Samme år ble Roger Kornberg (sønn av Nobelprisvinneren A. Kornberg) hedret med Nobelprisen i kjemi for sine studier om RNA -polymerase.
- I 2009 ble Ada Yonath tildelt Nobelprisen i kjemi "for studier om ribosomets struktur og funksjon" sammen med Venkatraman Ramakrishnan og Thomas A. Steitz .
RNA -rensing og deteksjon
RNA kan renses ved RNA -rensing , f.eks. B. ved RNA -ekstraksjon , skilles fra andre biomolekyler . Mengden og renheten av det isolerte RNA bestemmes ved fotometrisk måling ved en bølgelengde på 260 og 280 nm. Ytterligere informasjon om kvaliteten på RNA oppnås ved agarosegelelektroforese etterfulgt av farging med fargestoffer som SYBR Green II , metylenblått , Flekker-alt eller med en sølvfarging . Den kvalitative påvisningen av RNA (enten et bestemt RNA er tilstede) utføres for det meste ved RT-PCR , noen ganger med påfølgende DNA-sekvensering , eller ved Northern blot . Den kvantitative påvisningen (hvor mye av et bestemt RNA er tilstede) utføres for det meste med qRT-PCR , i tilfelle av rensede prøver med bare én RNA-sekvens kan konsentrasjonen også bestemmes ved fotometri . RNA kan strekkes og justeres gjennom molekylær kombinasjon . Ved bruk av in situ hybridisering kan spesifikke RNA påvises i celler og vev uten forutgående isolasjon.
bruk
RNA brukes til forskjellige formål. Når det gjelder ribozymer , har RNA en enzymatisk aktivitet , mens aptamerer inngår en langsiktig binding til en målstruktur. Kort dobbeltstrenget RNA i form av siRNA og shRNA brukes til midlertidig å undertrykke genuttrykk via RNA-interferens . RNA -vaksiner er en av de genetiske vaksinene der antigenet lages i cellene til personen som blir vaksinert. Noen CRISPR -Cas -systemer kan brukes til mer midlertidige redigeringer enn med DNA for å modifisere RNA - for eksempel for å behandle sykdommer. [6] En gratis plattform for design av RNA -målsekvenser ble publisert i 2020. [7] [8]
Se også
- Biontech
- Moderna
litteratur
- J. Marx: P-Bodies Marker stedet for kontroll av proteinproduksjon . I: Vitenskap . Bind 310, nr. 5749, s. 764-765. 2005, PMID 16272094 doi: 10.1126 / science.310.5749.764 .
- Seyffert: Lærebok i genetikk . 2. utgave, s. 42. Spektrum Akademischer Verlag, 2003.
- Albert Gossauer: Struktur og reaktivitet av biomolekyler - En introduksjon til organisk kjemi . S. 525. Wiley-VCH, 2006.
- Kapranov P, St Laurent G, Raz T, et al. : Flertallet av det totale atomkodede ikke-ribosomale RNA i en menneskelig celle er ikke-kommentert RNA av 'mørkt materie' . I: BMC Biol . 8, nr. 1, desember 2010, s. 149. doi : 10.1186 / 1741-7007-8-149 . PMID 21176148 .
Individuelle bevis
- ↑ Far A, Klussmann S: Slå speilbilde oligonukleotider inn i narkotika: utviklingen av Spiegelmer terapeutika. I: Drug Discovery Today . 20, nr. 1, januar 2015, s. 147–155. doi : 10.1016 / j.drudis.2014.09.004 . PMID 25236655 .
- ↑ Brosius, J. & Tiedge, H. (2004): RNomenclature. I: RNA Biol. 1 (2): 81-83. PMID 17179746 PDF
- ↑ S. Memczak, M. Jens inkluderer: Sirkulære RNA er en stor klasse av animalsk RNA med regulatorisk potens. I: Naturen . Bind 495, nummer 7441, mars 2013, s. 333–338, doi: 10.1038 / nature11928 . PMID 23446348 .
- ↑ Jeffrey M. Perkel: Besøker “Noncodarnia” . I: BioTechniques . 54, nr. 6, 2013, ISSN 1940-9818 . doi : 10.2144 / 000114037 .
- ↑ Anita G. Seto, Robert E. Kingston, Nelson C. Lau: The Coming of Age for Piwi Proteins . I: Molecular Cell . 26, nr. 5, 2007, ISSN 1097-2765 , s. 603-609. doi : 10.1016 / j.molcel.2007.05.021 .
- ^ Sara Reardon: Step Aside, CRISPR: RNA -redigering tar av ( en ) In: Scientific American . Hentet 25. september 2020.
- ↑ Ny type CRISPR-teknologi for å målrette mot RNA, inkludert RNA-virus som koronavirus (en-us) . I: phys.org . Hentet 3. april 2020.
- ^ Hans-Hermann Wessels, Alejandro Méndez-Mancilla, Xinyi Guo, Mateusz Legut, Zharko Daniloski, Neville E. Sanjana: Massivt parallelle Cas13-skjermer avslører prinsipper for veiledende RNA-design . I: Nature Biotechnology . 16. mars 2020, s. 1–6. doi : 10.1038 / s41587-020-0456-9 . PMC 7294996 (gratis fulltekst).
weblenker
- Litteratur av og om ribonukleinsyre i katalogen til det tyske nasjonalbiblioteket
- Thomas Tuschl - på vei til Nobelprisen ( Memento fra 7. desember 2007 i Internettarkivet ) Thomas Tuschl på stern.de om RNA -interferens, 19. juli 2004
- Die Zeit: Treasure in the genom , Die Zeit No. 48/2004 av 18. november 2004
- RNA